J Cardiovasc Thorac Res. 15(3):138-144.
doi: 10.34172/jcvtr.2023.31724
Original Article
Sevoflurane-mediated modulation of oxidative myocardial injury
Siavash Sedghi Writing – original draft, Writing – review & editing, 1, 2 
Wiam Z. Khadra Conceptualization, Methodology, Validation, 1, 2
Leili Pourafkari Conceptualization, Validation, Writing – review & editing, 1, 3
Paul R. Knight Conceptualization, Writing – review & editing, 1
Faraz A. Alderson Conceptualization, Data curation, Methodology, Validation, 4
Nader D. Nader Conceptualization, Data curation, Formal analysis, Project administration, Supervision, Validation, Visualization, Writing – review & editing, 1, 2, *
Author information:
1Department of Anesthesiology, University at Buffalo, Jacobs School of Medicine and Biomedical Sciences, Buffalo, New York, USA
2VA Western New York Healthcare System, Buffalo, New York, USA
3Cardiac Imaging, The Lundquist Institute, Harbor-University of California at Los Angles Medical Center, Los Angles, USA
4Faculty of Engineering, University of Guelph, Ontario, Canada
Abstract
Introduction:
Volatile anesthetics offer protection when administered throughout an ischemic injury. We examined how volatile anesthetics modulate the cardiac myocytic injury associated with hydrogen peroxide.
Methods:
Forty-eight Long-Evans rats were divided into four groups depending on the treatment: none (CONT), Glibenclamide (GLB); Sevoflurane (SEV); or GLB+SEV. Each group was further divided into two, one of which was exposed to hydrogen peroxide (H2O2). Oral GLB was administered 48 hours before myocardial isolation. All rats were anesthetized by intraperitoneal injection of Ketamine, and the hearts were harvested after heparinization. Cardiomyocytes were isolated using a combination of mechanical mincing and enzymatic digestion. After isolation, the aliquots of cells were exposed to H2O2 and FeSO4 for 30 minutes. The cell suspensions were then bubbled for 10 minutes with 100% oxygen and 1.5% SEV if appropriate. Apoptosis was detected by fluorescein-bound annexin-V (ANX-V), necrosis by propidium iodide, and ELISA assessed caspase-3 activity in all groups.
Results:
There was an increase in apoptosis, necrosis, and caspase-3 activity in the cells following exposure to hydrogen peroxide. SEV reduced the rate of cell necrosis and apoptosis. Pretreatment with GLB did not alter the effects of SEV. Similarly, caspase-3 activity did not change with GLB, although SEV administration reduced this enzymatic activity in response to hydrogen peroxide.
Conclusion:
In this oxidant injury model, we demonstrated that incubating isolated cardiomyocytes with SEV profoundly diminished H2O2-induced apoptotic and necrotic cells compared to their CONTs. These results support the hypothesis that KATP channels are not the sole mediators associated with anesthetic preconditioning.
Keywords: Apoptosis, Necrosis, Reactive oxygen species, Volatile anesthetics
Copyright and License Information
© 2023 The Author(s)
This is an open access article distributed under the terms of the Creative Commons Attribution License (
http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Introduction
Reactive oxygen species (ROS) are integral to many inflammatory and noninflammatory diseases. Although they also play a role in the normal homeostasis of the cell, an overabundance of ROS leads to a robust inflammatory response which directly or indirectly leads to cell death. Inflammatory responses associated with ROS have been well-studied in diseases such as cancer, rheumatic and neurodegenerative diseases, diabetes mellitus, and ischemia-reperfusion injuries.1,2 The importance of the injurious effects of ROS can be demonstrated in ischemic organs, such as the heart, following an acute myocardial infarction. For example, during myocardial infarction, the central nidus of the ischemic myocardium becomes necrotic and will later heal through scar formation. The myocardium adjacent to the infarct (the penumbra zone) following ischemia is functionally impaired (stunned) even though the myocardium may still be viable. Depending on the extent of ROS present, this penumbra zone may return to its normal function or become apoptotic, ultimately leading to heart failure.
It has been well-established that volatile anesthetics are potent immune modulators. They have decreased inflammatory responses, leading to cell death by either apoptosis or necrosis. The exact mechanism of action has yet to be well understood, but ATP-gated potassium (KATP) channels have been primarily implicated. Our team has previously demonstrated decreased inflammatory-related injury associated with ischemia-reperfusion injury in cardiopulmonary bypass patients.3
Our objective is to examine how volatile anesthetics modulate ROS-associated cellular injury. Based on our previous experimental and clinical findings, we hypothesize that volatile anesthetics have protective effects against apoptosis and necrosis in cardiomyocytes. We further hypothesize that this protection is primarily mediated through the activation and opening of KATP channels. To test this hypothesis, we will examine apoptosis and necrosis in isolated cardiomyocytes with inactivated KATP channels.
Materials and Methods
Experimental design
Forty-eight healthy male Long-Evans rats (Harlan-Sprague Dawley, Indianapolis, IN) with body weights of 250-300g were randomly divided into four different treatment groups: no treatment (CONT); Glibenclamide (GLB); Sevoflurane (SEV); or GLB with SEV. Each of these four treatment groups was randomly divided into two, one exposed to hydrogen peroxide (H2O2) (Table 1). Forty-eight hours before the experiment, all treatment groups involving GLB (groups 3, 4, 7, and 8) were treated once with 10mg/kg of GLB (Sigma Chemical Company, St. Louis, MO, United States) to induce deactivation of KATP channels. The treatment was administered through an 18-gauge gavage feeding tube about three inches long to ensure proper administration.
Table 1.
Treatment and intervention groups
|
Number
|
Group Name
|
Number of animals
|
| Group 1 |
No insult Controls (Negative Controls) |
6 |
| Group 2 |
Oxidative insult Controls (Positive Controls) |
6 |
| Group 3 |
Glibenclamide without Oxidative insult (Treatment Controls) |
6 |
| Group 4 |
Glibenclamide with Oxidative insult |
6 |
| Group 5 |
Sevoflurane without insult |
6 |
| Group 6 |
Sevoflurane with Oxidative insult |
6 |
| Group 7 |
Glibenclamide + Sevoflurane without insult |
6 |
| Group 8 |
Glibenclamide + Sevoflurane with Oxidative insult |
6 |
Anesthesia and myocardial tissue harvest cardiomyocyte isolation (Enzymatic Digestion)
The non-SEV groups (1-4) were anesthetized using ketamine (25 mg/kg) intraperitoneally (IP). The remainder of the groups (5-8) were anesthetized with SEV until animals had no pedal reflex after a firm toe pinch. SEV vapor (2%) with oxygen was delivered through a nose cone to maintain the anesthesia in groups 5 through 8. For tissue harvest, animals were heparinized (10,000 U/kg) (JA Webster, MA) with an injection into the ear vein to prevent clotting in the coronary arteries. Medial sternotomy was made by exposing the heart and ascending aorta by dissection of the pericardium. The heart was lifted anteriorly, exposing the pulmonary artery and the ascending aorta. The heart was isolated by transecting the proximal ascending aorta, leaving enough arterial stalk attached (proximal to the inanimate artery) to allow cannula placement without the aortic root. The heart was then removed and placed in a cold cardioplegic solution (modified Krebs-Hensleit solution) and was massaged to remove access blood from the chambers and the vasculature system before weighing by dripping a Krebs–Hensleit (KH) solution through the aorta.
The cells were then enzymatically broken up by breaking the calcium (Ca2+) bonds, subsequently digesting collagen and other connective tissues (KH solution without and with CaCl2 / collagenase, respectively). The heart was then mechanically diced into small pieces and incubated with KH solution with trypsin to break extracellular proteins and cleavage of gap junctions without disturbing the cells’ integrity. Trypsin activity was then stopped by adding Ca2+ back into the system. After a manual cell count, 250 000 cells were washed and resuspended in 5 milliliters of sterile KH solution.
Post-Isolation Treatment and Oxidative Injury
The CONT treatment groups (1 and 2) were exposed to 100% O2 for 30 minutes after isolation of the individual left ventricular myocytes. This was done by bubbling oxygen gas into the KH solution containing said cells at a flow rate of 400 mL/minute. Similarly, SEV-treated groups (5-8) were exposed to SEV at 2% and oxygen at 98% for 30 minutes using the same method. All cells were incubated in KH solution (pH = 7.4) at 37°C with 95% humidity and aerated with carbogen (O2 95% + CO2 5%).
Fenton reaction was utilized to induce oxidative damage to the cardiomyocytes by adding H2O2 and FeSO4 (groups 2, 4, 6, and 8). Cell suspensions were treated with 100μM H2O2 (Sigma Chemical Company, St. Louis MO, United States) and 200μM anhydrous FeSO4 (Sigma Chemical Company, St. Louis MO, United States). To stop the Fenton reaction, a mixture of 2 millimolar concentration (mM) of sodium etidronate (Sigma Chemical Company, St. Louis MO, United States) and 1mM sodium thiosulfate (Sigma Chemical Company, St. Louis MO, United States) in phosphate-buffered saline (PBS) (Burlington, MA, United States) were then added to the cell solutions. The cells were then subsequently washed with PBS.
Determination of Apoptosis / Necrosis
Annexin-5 (ANX-V) is a 35-36 kDa Ca2+ -dependent phospholipid-binding protein with a high affinity for phosphatidylserine (PS). Cells undergoing apoptosis have PS, usually on the inner leaflet translocated to the outer layer in the early apoptosis.4,5 ANX-V may be conjugated to a fluorochrome, including fluorescein isothiocyanate, thus detected by flow cytometry. Propidium iodide (PI) is used in conjunction with ANX-V on cells with a cell permeability (i.e., cells that have undergone necrosis).4 The nucleic acid intercalation bound to PI can also be visualized under flow cytometry (Figure 1).
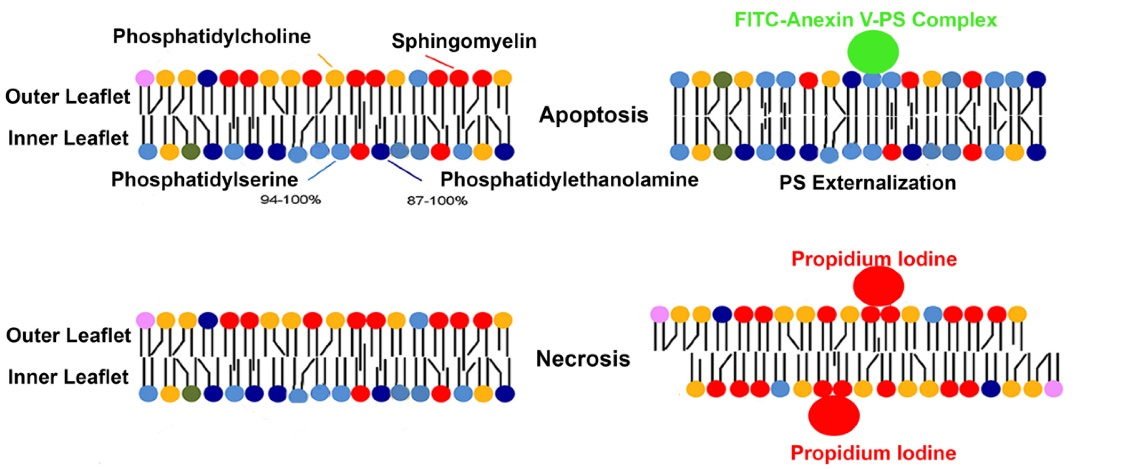
Figure 1.
The mechanism of fluorescence of Annexin-fluorescein isothiocyanate (FITC) and Propidium iodide (PI)
.
The mechanism of fluorescence of Annexin-fluorescein isothiocyanate (FITC) and Propidium iodide (PI)
A commercially available fluorescein isothiocyanate ANX-V Apoptosis detection Kit (Pharminogen, United States) was used to distinguish between apoptotic and necrotic cells. Briefly, the cells were washed twice with cold PBS and resuspended. 5 µl of fluorescein isothiocyanate - ANX-V and 5 µl PI were added to 100 000 cells. The cells were then gently vortexed and incubated for 15 minutes at room temperature (25°C) in the dark, and a binding buffer was added and analyzed using flow cytometry.
Caspase 3 assay
Caspases are crucial mediators of apoptosis, among which, Caspase 3 is an activated protease.6-8 To test for caspase 3 (a common mediator of the apoptotic pathway), we used a commercially available Caspase-3 Assay Kit (Abcam, Cambridge, UK). Briefly isolated left ventricular myocytes from all groups were lysed and incubated with DEVD-AFC (substrate for activated caspase-3) for 2 hours. The fluorescence intensities of the treated samples were compared using a Bio-Rad plate reader.
Data Management and Statistical Analysis
Six rats were studied in each group. The number of rats in each group was selected based on power analysis accepting a 20% beta error and 5% alpha error. Data were expressed as mean ± SEM and analyzed using a one-way analysis of variance with the Bonferroni post hoc test. Student’s t-tests were used for intergroup comparisons. Bonferroni correction was utilized for multiple comparisons and intergroup analyses. Null hypotheses were rejected at P value < 0.05.
Results
Gross apoptotic activity: An increase in necrosis and apoptosis is appreciated when isolated left ventricular myocytes are exposed to oxidative injury (Figure 2B) compared to CONT (Figure 2A). Treating cells with SEV decreased the observed injury (Figure 2C). A closer look at apoptotic activity amongst all non-ROS exposed cells (Figure 3) demonstrates that a larger increase in apoptosis of the myocytes was observed following ischemia in the CONT and GLB treated groups (9.6 ± 1.4% and 9.9 ± 1.5%, respectively with P value < 0.001). The difference between the CONT group and the myocytes that received GLB was negligible (P value > 0.999). Under the same oxidative insult, treatment with SEV decreased the apoptotic rate in the myocytes appreciatively drastically (4.4 ± 1.2%) (P value < 0.001). Oxidative injury demonstrated a marginal increase in the apoptotic rate compared to its non-ROS-treated counterpart (P value = 0.613). Even when GLB was added to SEV under the same oxidative conditions, there was very minimal change in the apoptotic rate (4.1 ± 0.8%, P value > 0.999), and similar to SEV, only a marginal increase from its non-ROS treated counterpart (P value = 0.460). The degree of apoptosis was similar among all groups not exposed to ROS injury.
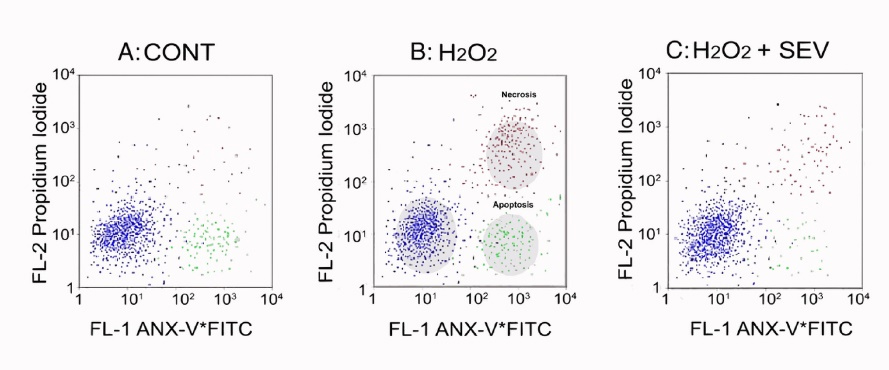
Figure 2.
The flow-cytometry comparison of Annexin-fluorescein isothiocyanate (ANX-V + FITC) and Propidium iodide (PI). 2A: Controls; 2B: Reactive oxygen species (ROS); and 2C: ROS with Sevoflurane (SEV). The cells’ characteristics are as follows: normal cells blue (lower left quadrant); apoptotic cells: ANX-V + FITC positive cells (FL-1: green; lower right quadrant), necrotic cells: PI-positive (FL-2: Red, upper right quadrant)
.
The flow-cytometry comparison of Annexin-fluorescein isothiocyanate (ANX-V + FITC) and Propidium iodide (PI). 2A: Controls; 2B: Reactive oxygen species (ROS); and 2C: ROS with Sevoflurane (SEV). The cells’ characteristics are as follows: normal cells blue (lower left quadrant); apoptotic cells: ANX-V + FITC positive cells (FL-1: green; lower right quadrant), necrotic cells: PI-positive (FL-2: Red, upper right quadrant)
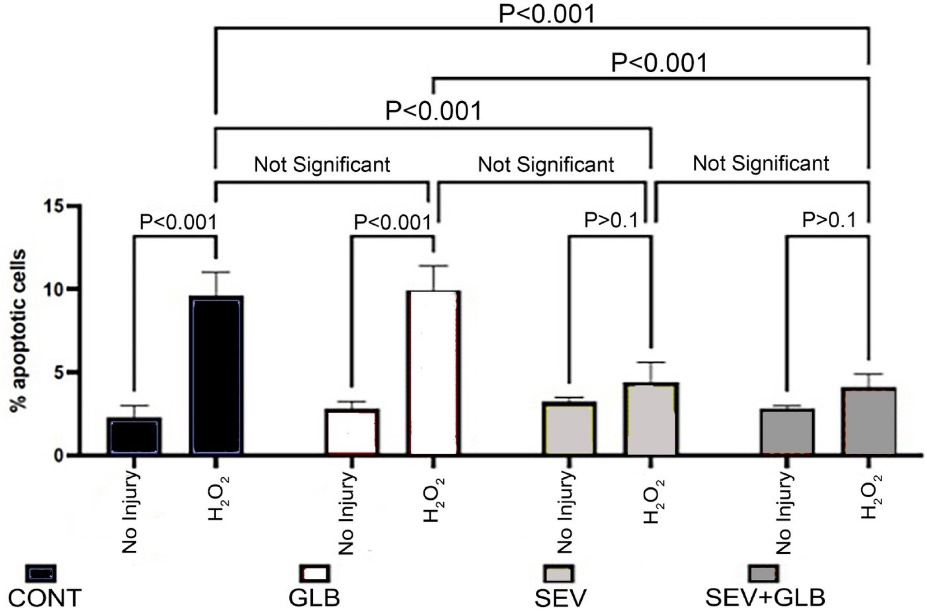
Figure 3.
The percentage of apoptotic cells two hours after various treatments. CONT: Controls; H2O2: Reactive oxygen species; GLB: Glibenclamide and SEV: Sevoflurane
.
The percentage of apoptotic cells two hours after various treatments. CONT: Controls; H2O2: Reactive oxygen species; GLB: Glibenclamide and SEV: Sevoflurane
Caspase 3 Activity
We also assessed increases in one of the chemical mediators of apoptosis, caspase 3 (Figure 4). The ROS-exposed myocytes in the CONT group had the highest mean caspase three fluorescence intensity (0.124 ± 0.009) compared to all groups under the same conditions (P value: < 0.001). This value decreased significantly when the ROS-exposed cells were treated with SEV (0.062 ± 0.006).
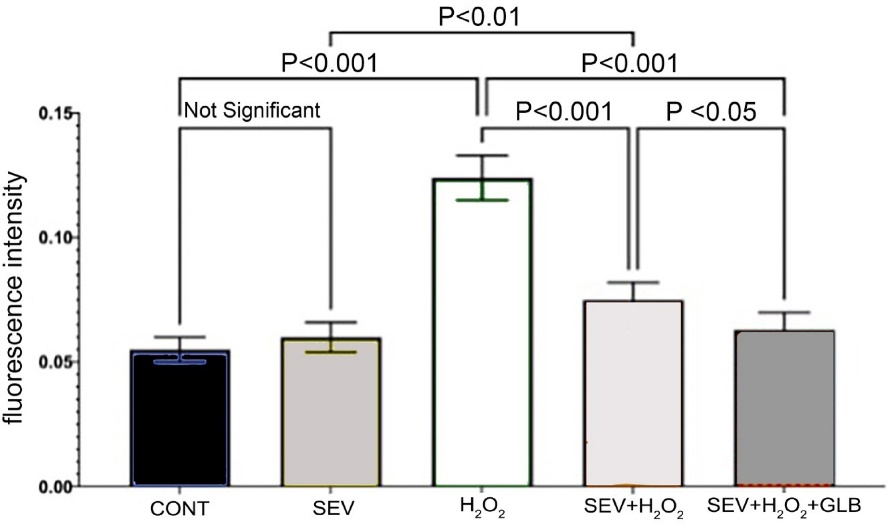
Figure 4.
Caspase-3 activity two hours after various treatments. CONT: Controls; H2O2: Reactive oxygen species; GLB: Glibenclamide and SEV: Sevoflurane
.
Caspase-3 activity two hours after various treatments. CONT: Controls; H2O2: Reactive oxygen species; GLB: Glibenclamide and SEV: Sevoflurane
Non-oxidant CONT (0.551 ± 0.005) and SEV-treated groups are comparable (P value = 0.758), serving as the CONTs. The no-treatment group had the highest level of caspase activity (0.124 ± 0.009). Treatment with SEV drastically reduced the mean caspase 3 activity (0.062 ± 0.006) (P value < 0.001). A similar effect was observed even in adding GLB to the SEV (0.063 ± 0.001; P value < 0.001). In oxidative injury, treatment with both GLB and SEV treatments further decreased caspase activity compared to SEV alone. Interestingly, comparing oxidative insult in myocytes exposed to both SEV + GLB was slightly lower than SEV under the same conditions (p value = 0.036).
Discussion
We have demonstrated that treating isolated cardiomyocytes with sevoflurane diminishes apoptotic and necrotic cell damage secondary to an oxidative injury. ROS provoke apoptotic injury mainly through upregulating pro-inflammatory genes.3,9-11 We previously demonstrated that sevoflurane decreases inflammatory responses after cardiopulmonary bypass by showing a functional improvement in the heart post-reperfusion.3
Reactive oxygen species cause necrotic and apoptotic cell damage through direct oxidizing effects on the macromolecules involved.12 This ROS-mediated damage underlies the pathogenesis of various diseases.13,14 For example, our laboratory demonstrated increased oxidant activity on lipids and proteins, causing lung injury following acid aspiration and high ambient oxygen concentrations.12 Periods of ischemia induce ROS in tissues triggering the activation of white blood cells (WBC). These activated WBCs can damage the involved tissues.15 Similarly, ROS-mediated apoptosis and necrosis of cardiomyocytes lead to heart failure following ischemia.16,17 In this study, we have shown that ROS leads to apoptosis and necrosis in isolated cardiomyocytes.
Apoptosis is an actively regulated process of cellular destruction without inflammation. Although the associated ischemic insult is less than is required for necrosis, apoptosis may still confer considerable dysfunction, as is observed in the myocardium. In an ischemic heart, necrosis is more prominent within the first 24 hours.16 However, the apoptotic changes present late after the ischemic insult due to reperfusion. This degree of myocardial injury may be reversible (“stunning”) or irreversible, leading to uncompensated heart dysfunction.16 On a molecular level, caspase 3 (a primary proteolytic mediator of apoptosis) directly correlates with the extent of cell death through autophagic processes. Caspases are involved in apoptotic cell death via proteolysis within the cells.6-8
KATP channels are strongly associated with apoptotic and necrotic cell injury. These channels open mainly during periods of energy depletion, as seen in ischemic events, thus mediating responses involved in stress adaptation.18,19 KATP channels are found on the cardiac myocytes sarcolemma and the mitochondrial membrane.19,20 Volatile anesthetics increase the opening of KATP channels leading to a decrease in the cytosolic and mitochondrial calcium load. This mechanism offers primary protection against calcium accumulation and mitochondrial dysfunction, as is seen with ischemia-reperfusion injury.21,22 Opening the KATP channels decreases the calcium influx and restores the mitochondrial membrane potential, thus preventing apoptosis. Opening further leads to an increased ATP synthesis which protects against cellular injury.23,24 The sarcolemmal KATP channels decrease the O2 and ATP consumption and may shorten the action potential, leading to myocardial protection.23
Deactivating KATP channels with pharmacological inhibitors such as GLB hinders or diminishes ischemic and anesthetic preconditioning protection.25-27 Interestingly, unlike these studies with the deactivation of KATP channels, we demonstrated only minor attenuation of anesthetic preconditioning with SEV. Due to the complexity of anesthetic preconditioning, the causality of our contradicting results still needs to be fully understood. We hypothesize that KATP channels are not our model’s sole mediators associated with anesthetic protection.
Mitochondrial permeability transition pores (MPTP) also play an essential role in apoptosis during a sustained injury. When these channels are in an irreversible open state, there is a disruption of the mitochondrial potential and subsequent swelling of the mitochondria. With the swelling of mitochondria, pro-apoptotic proteins such as cytochrome c are released into the cytosol triggering a cascade of events that lead to apoptosis or necrosis.28
Similar to KATP channels, MPTPs are vital in myocardial preconditioning. Volatile anesthetics delay the opening of these channels under oxidative stress in rat cardiomyocytes and cardiomyocyte-derived embryonic stem cells, which leads to cellular protection.29-33 It is hypothesized that the incorporation of lipid-soluble volatile anesthetics reversibly into the lipid bilayer component of the cell membrane causes a lateral pressure favoring the closed state of the MPTPs, which may contribute to ischemia protection.34
Additionally, volatile anesthetics may directly modulate anti-apoptotic genes. Preconditioning with volatile anesthetics for one hour provides neuroprotection by upregulating the anti-apoptotic genes.35 In rat neurons exposed to volatile anesthetics, an increased ratio of the anti-apoptotic Bcl-2 protein family to a decreased pro-apoptotic activation of the c-Jun N-terminal kinase (JNK) and p53 pathway led to a decrease in caspase-3 expression.36 The tandem of P-domains in weakly inward rectifying K+ channels (TWIK) is another set of ion channels associated with apoptotic cell death. TWIK-related acid-sensitive potassium channels (TASK) 1 through 3 inhibit the intracellular apoptotic pathways and thus enhance cellular viability. Volatile anesthetics augment these protective effects by enhancing the TASK3 activity.37,38
Limitations in our study may include alterations induced by isolating cardiomyocytes on the pathways involved in apoptosis, such as that of KATP channels. The isolation of the cells may alter many ways involved in physiological functions. Furthermore, the timing and dosage of the GLB may have been insufficient to deactivate the KATP channels in this model. Additionally, we have considered whether GLB should have been given post-isolation to ensure a compatible ratio of SEV and GLB in the system.
Conclusion
In a previous review article, we have outlined the immune modulatory effects of the volatiles anesthetics.39 Along with the many other studies conducted so far, only future studies and experimental work can answer the question of the different pathways associated with anesthetic preconditioning responsible for our findings. To investigate the direct effects of MPTPs on anesthetic preconditioning, the blockage of both KATP channels with the MPTPs may clarify the mechanism responsible for our results. Finally, to better understand the cellular mechanisms of volatile anesthetic action, hyperbaric pressure experiments may be promising as high atmospheric pressures are shown to reverse many volatile anesthetics actions.40
Competing Interests
None declared by the authors.
Ethical Approval
All experimental procedures and protocols used in this study were reviewed and approved by the Animal Use and Care Committee of the VA Medical Center at Buffalo (Approval letter No: 55443-16326 dated Feb 18, 2014), conforming to the Guiding Principles in the Care and Use of Animals of the American Physiologic Society.
Funding
Funding was provided for this study by the grant-in-aid from the American Heart Association (00604240) and the Society of Cardiothoracic Anesthesiology awarded to NDN.
References
- Hori M, Nishida K. Oxidative stress and left ventricular remodelling after myocardial infarction. Cardiovasc Res 2009; 81(3):457-64. doi: 10.1093/cvr/cvn335 [Crossref] [ Google Scholar]
- Yang S, Lian G. ROS and diseases: role in metabolism and energy supply. Mol Cell Biochem 2020; 467(1-2):1-12. doi: 10.1007/s11010-019-03667-9 [Crossref] [ Google Scholar]
- Nader ND, Li CM, Khadra WZ, Reedy R, Panos AL. Anesthetic myocardial protection with sevoflurane. J Cardiothorac Vasc Anesth 2004; 18(3):269-74. doi: 10.1053/j.jvca.2004.03.004 [Crossref] [ Google Scholar]
- Crowley LC, Marfell BJ, Scott AP, Waterhouse NJ. Quantitation of apoptosis and necrosis by annexin V binding, propidium iodide uptake, and flow cytometry. Cold Spring Harb Protoc 2016; 2016(11):953-7. doi: 10.1101/pdb.prot087288 [Crossref] [ Google Scholar]
- van Heerde WL, Robert-Offerman S, Dumont E, Hofstra L, Doevendans PA, Smits JF. Markers of apoptosis in cardiovascular tissues: focus on annexin V. Cardiovasc Res 2000; 45(3):549-59. doi: 10.1016/s0008-6363(99)00396-x [Crossref] [ Google Scholar]
- Black SC, Huang JQ, Rezaiefar P, Radinovic S, Eberhart A, Nicholson DW. Co-localization of the cysteine protease caspase-3 with apoptotic myocytes after in vivo myocardial ischemia and reperfusion in the rat. J Mol Cell Cardiol 1998; 30(4):733-42. doi: 10.1006/jmcc.1998.0660 [Crossref] [ Google Scholar]
- Porter AG, Jänicke RU. Emerging roles of caspase-3 in apoptosis. Cell Death Differ 1999; 6(2):99-104. doi: 10.1038/sj.cdd.4400476 [Crossref] [ Google Scholar]
- Wu MY, Yiang GT, Liao WT, Tsai AP, Cheng YL, Cheng PW. Current mechanistic concepts in ischemia and reperfusion injury. Cell Physiol Biochem 2018; 46(4):1650-67. doi: 10.1159/000489241 [Crossref] [ Google Scholar]
- Lingappan K. NF-κB in oxidative stress. Curr Opin Toxicol 2018; 7:81-6. doi: 10.1016/j.cotox.2017.11.002 [Crossref] [ Google Scholar]
- Mittal M, Siddiqui MR, Tran K, Reddy SP, Malik AB. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal 2014; 20(7):1126-67. doi: 10.1089/ars.2012.5149 [Crossref] [ Google Scholar]
- Wu J, Cai W, Du R, Li H, Wang B, Zhou Y. Sevoflurane alleviates myocardial ischemia reperfusion injury by inhibiting P2X7-NLRP3 mediated pyroptosis. Front Mol Biosci 2021; 8:768594. doi: 10.3389/fmolb.2021.768594 [Crossref] [ Google Scholar]
- Nader-Djalal N, Knight PR 3rd, Thusu K, Davidson BA, Holm BA, Johnson KJ. Reactive oxygen species contribute to oxygen-related lung injury after acid aspiration. Anesth Analg 1998; 87(1):127-33. doi: 10.1097/00000539-199807000-00028 [Crossref] [ Google Scholar]
- Moris D, Spartalis M, Spartalis E, Karachaliou GS, Karaolanis GI, Tsourouflis G. The role of reactive oxygen species in the pathophysiology of cardiovascular diseases and the clinical significance of myocardial redox. Ann Transl Med 2017; 5(16):326. doi: 10.21037/atm.2017.06.27 [Crossref] [ Google Scholar]
- Zhao J, Wang F, Zhang Y, Jiao L, Lau WB, Wang L. Sevoflurane preconditioning attenuates myocardial ischemia/reperfusion injury via caveolin-3-dependent cyclooxygenase-2 inhibition. Circulation 2013; 128(11 Suppl 1):S121-9. doi: 10.1161/circulationaha.112.000045 [Crossref] [ Google Scholar]
- Zhou T, Prather ER, Garrison DE, Zuo L. Interplay between ROS and antioxidants during ischemia-reperfusion injuries in cardiac and skeletal muscle. Int J Mol Sci 2018; 19(2):417. doi: 10.3390/ijms19020417 [Crossref] [ Google Scholar]
- Garg S, Narula J, Chandrashekhar Y. Apoptosis and heart failure: clinical relevance and therapeutic target. J Mol Cell Cardiol 2005; 38(1):73-9. doi: 10.1016/j.yjmcc.2004.11.006 [Crossref] [ Google Scholar]
- Kametsu Y, Osuga S, Hakim AM. Apoptosis occurs in the penumbra zone during short-duration focal ischemia in the rat. J Cereb Blood Flow Metab 2003; 23(4):416-22. doi: 10.1097/01.wcb.0000052281.23292.7c [Crossref] [ Google Scholar]
- Lefer DJ, Nichols CG, Coetzee WA. Sulfonylurea receptor 1 subunits of ATP-sensitive potassium channels and myocardial ischemia/reperfusion injury. Trends Cardiovasc Med 2009; 19(2):61-7. doi: 10.1016/j.tcm.2009.04.008 [Crossref] [ Google Scholar]
- O’Rourke B. Myocardial K(ATP) channels in preconditioning. Circ Res 2000; 87(10):845-55. doi: 10.1161/01.res.87.10.845 [Crossref] [ Google Scholar]
- Zhuo ML, Huang Y, Liu DP, Liang CC. KATP channel: relation with cell metabolism and role in the cardiovascular system. Int J Biochem Cell Biol 2005; 37(4):751-64. doi: 10.1016/j.biocel.2004.10.008 [Crossref] [ Google Scholar]
- Holmuhamedov EL, Wang L, Terzic A. ATP-sensitive K + channel openers prevent Ca2 + overload in rat cardiac mitochondria. J Physiol 1999; 519(Pt 2):347-60. doi: 10.1111/j.1469-7793.1999.0347m.x [Crossref] [ Google Scholar]
- Yellon DM, Downey JM. Preconditioning the myocardium: from cellular physiology to clinical cardiology. Physiol Rev 2003; 83(4):1113-51. doi: 10.1152/physrev.00009.2003 [Crossref] [ Google Scholar]
- Wang Y, Haider HK, Ahmad N, Ashraf M. Mechanisms by which K(ATP) channel openers produce acute and delayed cardioprotection. Vascul Pharmacol 2005; 42(5-6):253-64. doi: 10.1016/j.vph.2005.02.008 [Crossref] [ Google Scholar]
- Zhang S, Du X, Zhang K, Wang H. Effects of sevoflurane on apoptosis of myocardial cells in IRI rats. Biomed Res Int 2021; 2021:3347949. doi: 10.1155/2021/3347949 [Crossref] [ Google Scholar]
- De Hert SG, Turani F, Mathur S, Stowe DF. Cardioprotection with volatile anesthetics: mechanisms and clinical implications. Anesth Analg 2005; 100(6):1584-93. doi: 10.1213/01.ane.0000153483.61170.0c [Crossref] [ Google Scholar]
- Gross GJ, Auchampach JA. Blockade of ATP-sensitive potassium channels prevents myocardial preconditioning in dogs. Circ Res 1992; 70(2):223-33. doi: 10.1161/01.res.70.2.223 [Crossref] [ Google Scholar]
- Tomai F, Crea F, Gaspardone A, Versaci F, De Paulis R, Penta de Peppo A. Ischemic preconditioning during coronary angioplasty is prevented by glibenclamide, a selective ATP-sensitive K + channel blocker. Circulation 1994; 90(2):700-5. doi: 10.1161/01.cir.90.2.700 [Crossref] [ Google Scholar]
- Kwong JQ, Molkentin JD. Physiological and pathological roles of the mitochondrial permeability transition pore in the heart. Cell Metab 2015; 21(2):206-14. doi: 10.1016/j.cmet.2014.12.001 [Crossref] [ Google Scholar]
- Bu HM, Yang CY, Wang ML, Ma HJ, Sun H, Zhang Y. K(ATP) channels and MPTP are involved in the cardioprotection bestowed by chronic intermittent hypobaric hypoxia in the developing rat. J Physiol Sci 2015; 65(4):367-76. doi: 10.1007/s12576-015-0376-5 [Crossref] [ Google Scholar]
- Canfield SG, Sepac A, Sedlic F, Muravyeva MY, Bai X, Bosnjak ZJ. Marked hyperglycemia attenuates anesthetic preconditioning in human-induced pluripotent stem cell-derived cardiomyocytes. Anesthesiology 2012; 117(4):735-44. doi: 10.1097/ALN.0b013e3182655e96 [Crossref] [ Google Scholar]
- Onishi A, Miyamae M, Kaneda K, Kotani J, Figueredo VM. Direct evidence for inhibition of mitochondrial permeability transition pore opening by sevoflurane preconditioning in cardiomyocytes: comparison with cyclosporine A. Eur J Pharmacol 2012; 675(1-3):40-6. doi: 10.1016/j.ejphar.2011.11.040 [Crossref] [ Google Scholar]
- Pravdic D, Sedlic F, Mio Y, Vladic N, Bienengraeber M, Bosnjak ZJ. Anesthetic-induced preconditioning delays opening of mitochondrial permeability transition pore via protein kinase C-epsilon-mediated pathway. Anesthesiology 2009; 111(2):267-74. doi: 10.1097/ALN.0b013e3181a91957 [Crossref] [ Google Scholar]
- Sepac A, Sedlic F, Si-Tayeb K, Lough J, Duncan SA, Bienengraeber M. Isoflurane preconditioning elicits competent endogenous mechanisms of protection from oxidative stress in cardiomyocytes derived from human embryonic stem cells. Anesthesiology 2010; 113(4):906-16. doi: 10.1097/ALN.0b013e3181eff6b7 [Crossref] [ Google Scholar]
- Cantor RS. The lateral pressure profile in membranes: a physical mechanism of general anesthesia. Biochemistry 1997; 36(9):2339-44. doi: 10.1021/bi9627323 [Crossref] [ Google Scholar]
- Bedirli N, Bagriacik EU, Emmez H, Yilmaz G, Unal Y, Ozkose Z. Sevoflurane and isoflurane preconditioning provides neuroprotection by inhibition of apoptosis-related mRNA expression in a rat model of focal cerebral ischemia. J Neurosurg Anesthesiol 2012; 24(4):336-44. doi: 10.1097/ANA.0b013e318266791e [Crossref] [ Google Scholar]
- Wang H, Shi H, Yu Q, Chen J, Zhang F, Gao Y. Sevoflurane preconditioning confers neuroprotection via anti-apoptosis effects. Acta Neurochir Suppl 2016; 121:55-61. doi: 10.1007/978-3-319-18497-5_10 [Crossref] [ Google Scholar]
- Meadows HJ, Randall AD. Functional characterisation of human TASK-3, an acid-sensitive two-pore domain potassium channel. Neuropharmacology 2001; 40(4):551-9. doi: 10.1016/s0028-3908(00)00189-1 [Crossref] [ Google Scholar]
- Patel AJ, Lazdunski M. The 2P-domain K + channels: role in apoptosis and tumorigenesis. Pflugers Arch 2004; 448(3):261-73. doi: 10.1007/s00424-004-1255-8 [Crossref] [ Google Scholar]
- Sedghi S, Kutscher HL, Davidson BA, Knight PR. Volatile anesthetics and immunity. Immunol Invest 2017; 46(8):793-804. doi: 10.1080/08820139.2017.1373905 [Crossref] [ Google Scholar]
- Cohen PJ, Bedows E, Brabec MJ, Knight PR. Hyperbaric pressure of 51 atmospheres is without effect on the depression of oxygen uptake in kidney tissue culture produced by halothane. Life Sci 1985; 37(13):1221-4. doi: 10.1016/0024-3205(85)90133-x [Crossref] [ Google Scholar]